JR equations for psychologists
Nowadays, computational neuroscience brings together experts from a wide range of backgrounds: physicists, mathematicians, engineers, biologists and, lately, also psychologists like myself. There will always be room for questions such as “how did a psychologist end up doing this?”. My answer would be that I was looking for accuracy in how we understand cognition. The search led me to the brain, then to the crucial role of mathematics. Then, it came coding, and I love it.
In this post, I’d like to share my psychologist-friendly approach to expressing the Jansen-Rit equations. This formulation was inspired by the one used by Spiegler et al. (2010), and I used it in a recent eNeuro publication (Cabrera-Álvarez, 2024). It goes like this:
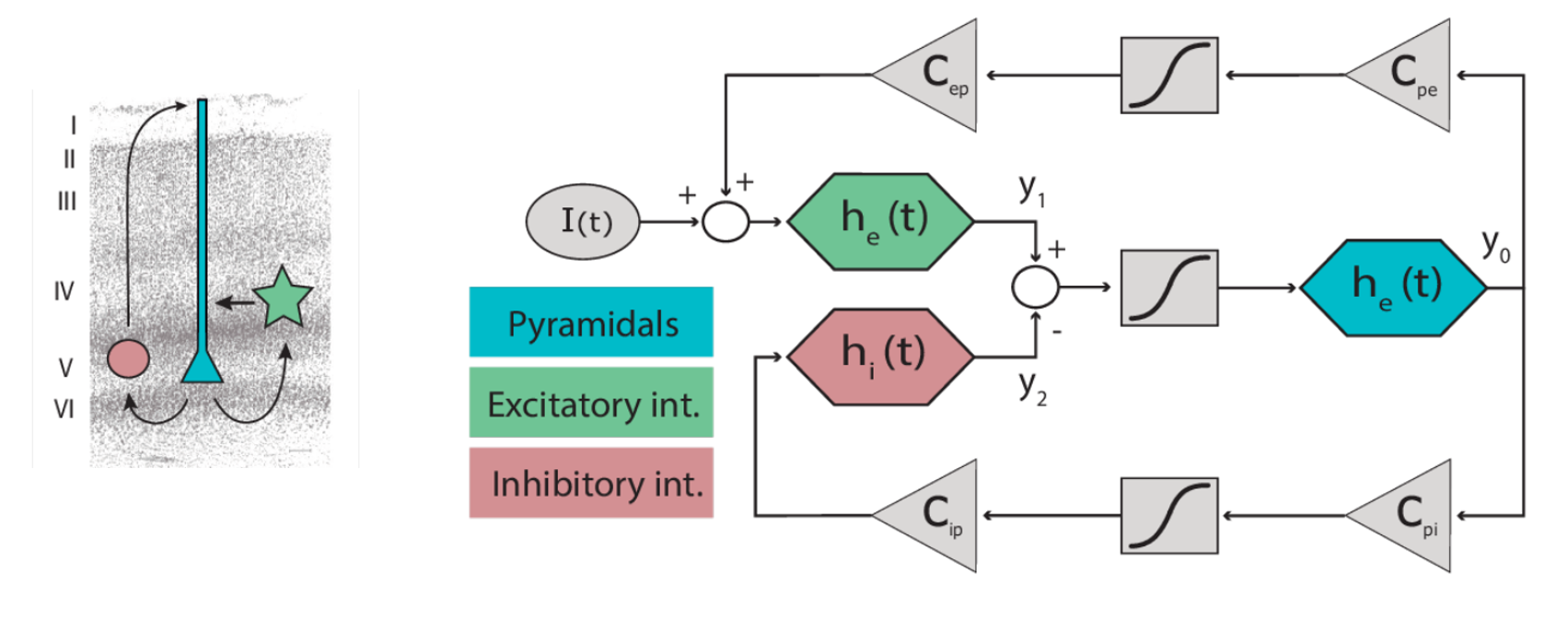
\[\begin{aligned} \dot y_{0_i} &= y_{3_i} \\ \dot y_{1_i} &= y_{4_i} \\ \dot y_{2_i} &= y_{5_i} \\ \dot y_{3_i} &= \frac{H_e}{\tau_e} \, S\left(y_{1_i} - y_{2_i}\right) - \frac{2}{\tau_e} \, y_{3_i} - \frac{1}{\tau_e^2} \, y_{0_i} \\ \dot y_{4_i} &= \frac{H_e}{\tau_e} \left[ I_i(t) + C_{ep} \, S(C_{pe} \, y_{0_i}) \right] - \frac{2}{\tau_e} \, y_{4_i} - \frac{1}{\tau_e^2} \, y_{1_i} \\ \dot y_{5_i} &= \frac{H_i}{\tau_i} \, C_{ip} \, S(C_{pi} \, y_{0_i}) - \frac{2}{\tau_i} \, y_{5_i} - \frac{1}{\tau_i^2} \, y_{2_i} \end{aligned}\]Post-synaptic potentials are now represented by $H$, referring to the impulse response function $h(t)$ that transforms firing rate into membrane voltage. Time constants are denoted by $\tau$, a conventional symbol used when describing synaptic dynamics. Additionally, the parameters governing synaptic connectivity are subscripted using a “from-to” convention—e.g., $C_{pe}$ indicates projections from pyramidal cells to excitatory interneurons. Take a look at the schematic below:
The sigmoidal activation function, the function that converts voltage into firing rate, did not change: \(S[v] = \frac{2 e_0}{1 + e^{r (v_0 - v)}}\)
And for the input to the system, we used $\eta$ for noise, representing $\mathcal{N}(p, \sigma)$ a white noise derived from a gaussian distribution with $p$ mean and $\sigma$ std. \(I_i(t) = \eta_i(t) + g \sum_{j=1}^{N} w_{ji} \, S\left[y_{1_j}(t - d_{ji}) - y_{2_j}(t - d_{ji})\right]\)
Conduction delays between regions is given by $d_{ji} = \frac{L_{ji}}{s}$.
Take a look to the parameters and their descriptions:
| Parameter | Value | Unit | Description |
|---|---|---|---|
| $H_e$ | 3.25 | mV | Average excitatory synaptic gain |
| $H_i$ | 22 | mV | Average inhibitory synaptic gain |
| $\tau_e$ | 10 | ms | Time constant of excitatory PSP |
| $\tau_i$ | 20 | ms | Time constant of inhibitory PSP |
| $C_{pe}$ | 135 | – | Synaptic contacts: pyramidals → excitatory interneurons |
| $C_{ep}$ | 108 | – | Synaptic contacts: excitatory interneurons → pyramidals |
| $C_{pi}$ | 33.75 | – | Synaptic contacts: pyramidals → inhibitory interneurons |
| $C_{ip}$ | 33.75 | – | Synaptic contacts: inhibitory interneurons → pyramidals |
| $e_0$ | 0.0025 | ms$^{-1}$ | Half of the maximum firing rate |
| $r$ | 0.56 | mV$^{-1}$ | Slope of the sigmoid at $v_0$ |
| $v_0$ | 6 | mV | Membrane potential for half-maximum firing rate |
| $p$ | 0.1085 | ms$^{-1}$ | Mean of intrinsic noisy input |
| $\sigma$ | 0.022 | ms$^{-1}$ | Standard deviation of intrinsic noise |
Enjoy Reading This Article?
Here are some more articles you might like to read next: